For the past year I’ve been working on a project involving the development of hair. Some of our recent findings involve variations in function of a key protein, a type of early developmental “morphogen” called Bone Morphogenetic Protein (BMP). For those not familiar, a morphogen is a class of molecule that helps to determine the shape of an organism or a given tissue type. So, many of the molecules involved in very early embryogenesis are considered morphogens, as you might imagine. In my work, we’ve been looking at variations from head-to-rump in patterns of BMP. And while I can’t say exactly what we’ve found (it’s still to be published!), I will say that there seem to be distinct differences in the roles BMP plays in the skin of the mouse from anterior (head) to posterior (rump), even though these skin and hair cells may look identical under the microscope. Chemically, they appear to be quite different, and therefore, as the title says, “cellular appearances can be deceiving”.

What this work has illustrated to me, alongside work from other investigators before me, is that even though different cells within a given tissue, such as the skin, may look identical, chemically and functionally they are unique. They are “differentiated” from their close relatives.
As an example taken from J. Michael Barry’s book, Molecular Embryology: How Molecules Give Birth to Animals, Barry talks about a series of experiments performed on the insect, Rhodinus.

Researchers performed various grafting experiments on Rhodinus embryos, removing sections of ectoderm and replacing them elsewhere on the body and studying different patterns of cellular growth in and around the tissue in the adult insect. Below is an illustration of some of those experiments (p. 74).
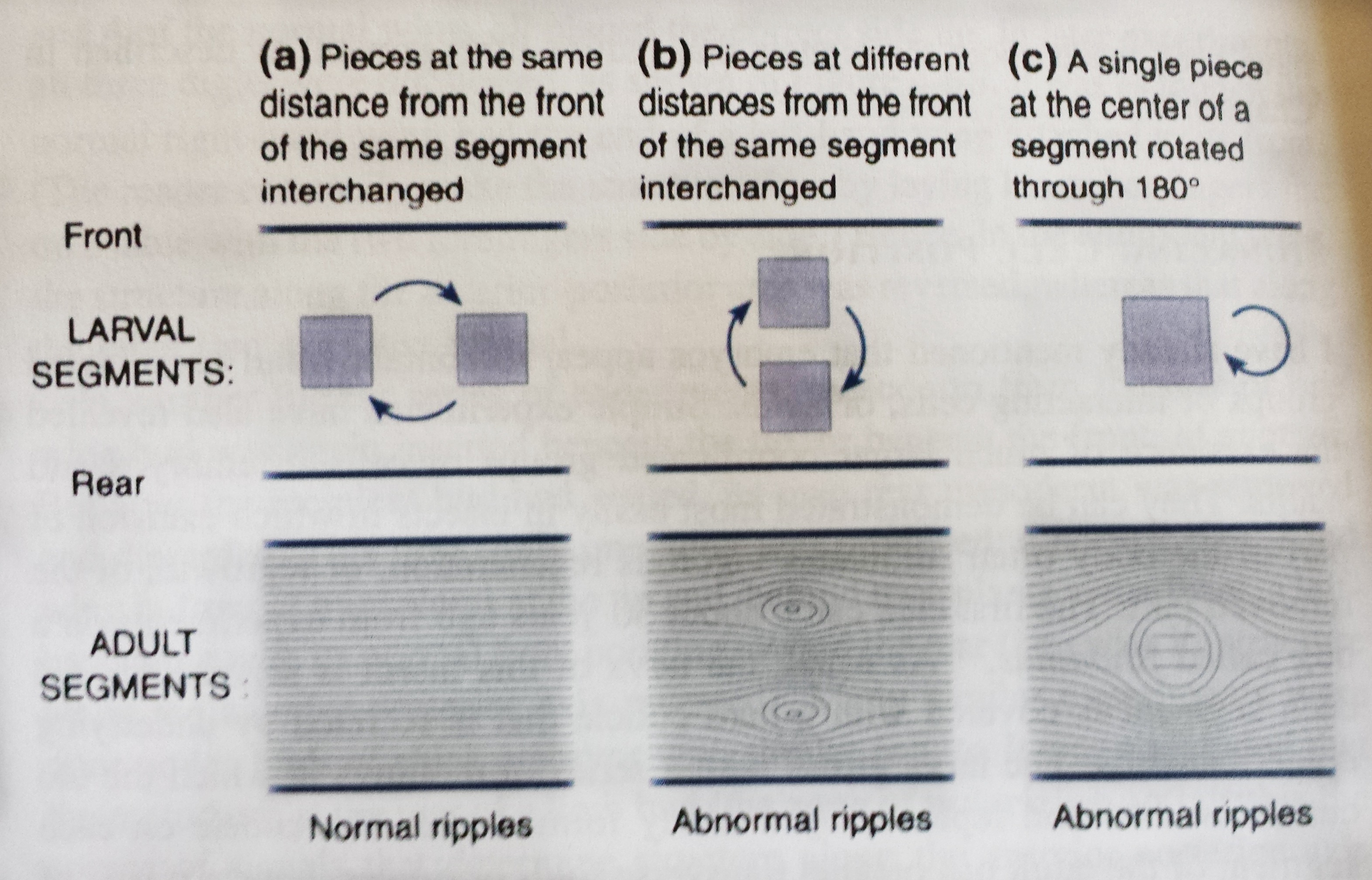
In (a) illustrated above, two squares of ectoderm from the same anterior-posterior plane in the body of the embryonic insect were switched, without any rotations to the tissue. In this instance, the tissue healed normally and looked relatively indiscernible from untreated insects. In illustration (b), however, the researchers took two segments of ectoderm from different planes within the anterior-posterior axis and switched them. In these insects, not only did healing occur, but the grafts caused an expansion of cell growth surrounding the segments. Likewise, as illustrated in (c), when a single segment of tissue ectoderm was rotated 180 degrees, there was an expansive growth surrounding the graft.
What this set of experiments suggests is that cells along the anterior-posterior (head-to-rump) axis, in some sense, “know” their locations and who their cellular neighbors are supposed to be. This is again illustrated in another set of experiments performed on cockroach embryos.
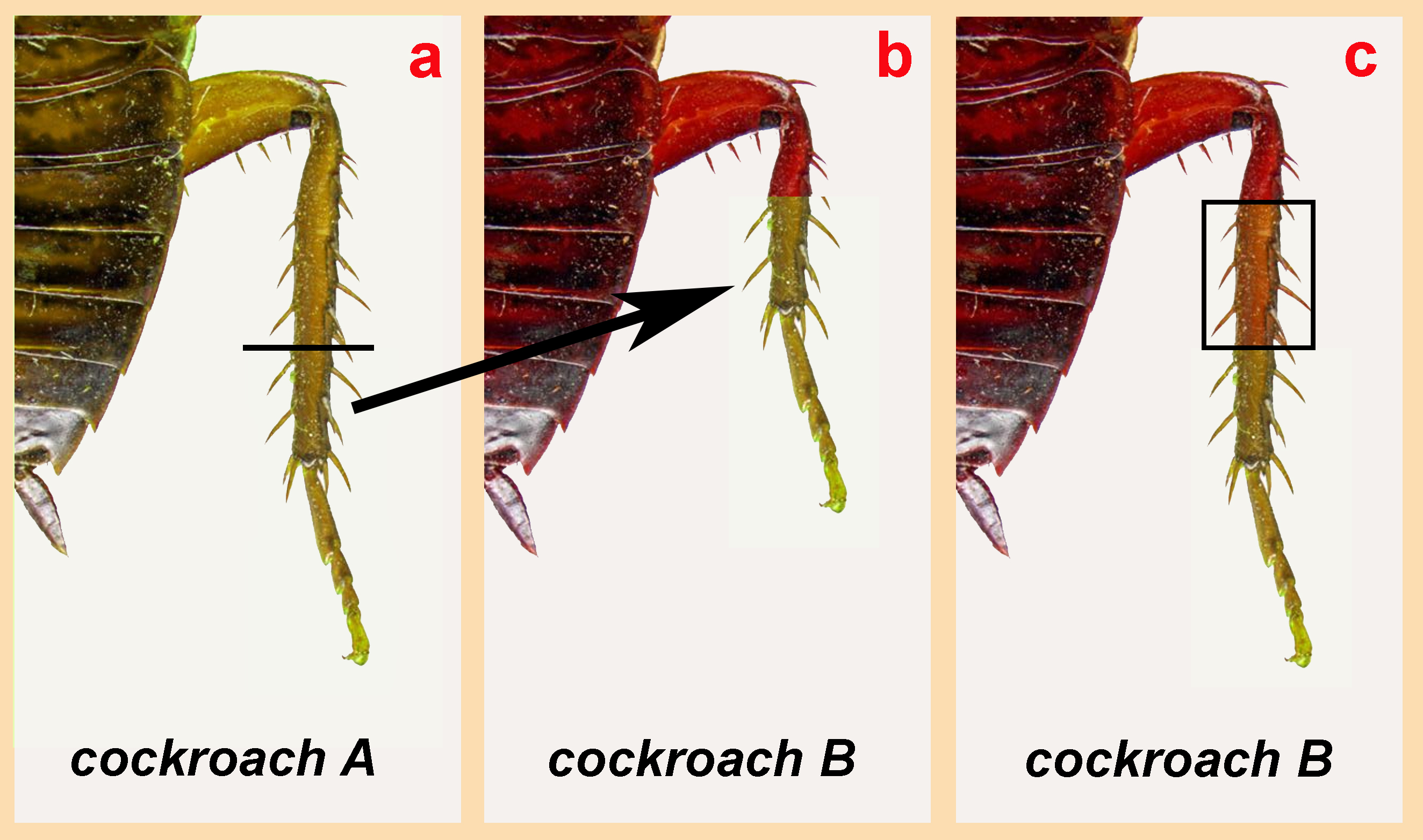
In this next set of experiments, illustrated by the image above, researchers took cockroach embryos and grafted legs from different cockroaches onto one another to see what would happen. In the image above, (a) shows a single cockroach “A” whose leg has been removed in embryo. Cockroach B has a portion of one of its own legs removed, but this surgery is performed further up along the leg than the portion removed from the donor. Following the graft of donor tissue (Cockroach A) onto Cockroach B’s leg, the B embryo has a stunted leg. By the time it’s an adult, however, not only has Cockroach B’s leg healed, it’s replaced the length of the leg that was originally missing after the graft (c). In short, the native tissue and the graft tissue both somehow knew that they weren’t supposed to be “neighbors” and reproduced the missing portion of the leg. It’s as if cells have surface molecules that tell them, in exceptional detail, who they’re saddled up next to. When they’re not next to the cells they’re supposed to be next to, this promotes tissue expansion in the attempt to regrow those cells which are missing. (Mind you, this is most readily apparent during the embryonic period; less so in adulthood.)
We scientists, and humans in general, really love our labels. In terms of the body, we label one thing a “brain”, another thing a “hand”, and yet another “skin”, when in fact, these are just gross generalizations of large multicellular tissues. We like to think of a tissue as relatively homogenous, and, yes, skin cells in different regions of the body probably have more in common with one another, molecularly, than, say, they do with the liver. But as my own experiments have highlighted for me, very subtle yet very important variations occur across cells that may share similar or even identical morphologies (shapes). In some sense, it may be better to consider each cell relatively unique and not get trapped into the mistake of conceptually homogenizing organs as though they’re some sort of cellular purée. Even though we may label the skin or the brain as singular organs, they are each made up of conglomerates of very different cells indeed.

Hi Emily,
fantastic. Thank you for your post. The morphogenetic hypothesis I shared with you some time ago could little be confirmed by all the experiments you reported: the adult individuals of any species already exist as electromagnetic-like molds, since fertilization during development until death.
“What this set of experiments suggests is that cells along the anterior-posterior (head-to-rump) axis, in some sense, “know” their locations and who their cellular neighbors are supposed to be. This is again illustrated in another set of experiments performed on cockroach embryos.”
The expression “in some sense” you use, by which cells “know” their locations, in my opinion is the sense +/- of the micro electromagnetic current of which living organisms are composed along with cellular matter. The micro electromagnetic mold forces the cells that are duplicating to occupy the right places and forms: as in the electromagnetic mold, so in the cellular copy (as in heaven, so on earth).
You can get the same results as in the experiments quoted by Barry, if you change a bit the same experiment I suppose you made when you were a child. Hold a magnet hidden under your desk surface. Then make some iron dust fall over the desk, you will see the dust assumes the form of the force lines of the magnet. When you withdraw the magnet, the iron dust continues to maintain the force lines shape until different forces will disarranged it. But, if no one come there to blow the dust away from your desk, you can “edit” some areas of the drawing, giving them different forms (square, circle etc.), and even replacing some iron dust area with rice grains.
Religions books in some case told scientific truths. I have readapted the prayer “Our Father” to modern scientific convenience.
http://www.meso.it/our-father-our-mold.htm
Warm regards,
Stefano
I can definitely see how attraction/repulsion works in the diffusion of molecules that form early gradients in embryo. For instance, related to my own experiments, diffusion of the protein, Wnt, has been described in this way. Also, membrane-bound chemotaxis-related molecules (attractants and repellents) are a popular area of study in the motility of cells. 🙂
“In some sense, it may be better to consider each cell relatively unique….”
I think that’s a key insight. So, I invite you to consider more carefully the aptness of qualifying the term “unique” by the adjective, “relatively.”
What if, in fact, each cell was–even if in “only” the most minute way–unique, different from every other cell, even of the same tissue type? What are the implications of such a view for biological processes? It might suggest that, while chemical in character, the processes which occur in cell repoduction never the less might, in the process of billions of iterations of the same cell types, produce minute differences in each cell which are hardly detectable by most ways of considering them.
I recall a fellow who states emphatically that, indeed, each cell is not merely relatively unique but, more simply, unique, however similar it may be to other cells of its type.
Hehe, good point. I suppose I used “relatively” to soften to idea. I do indeed believe every cell to be unique (just by its spatial location alone, that would lead to more or less minute chemical differences). Although, to consider every cell unique makes the building of biological models, well, a bit tricky. So taking into account “uniqueness” but also the larger system’s inter-relatedness, then perhaps some sort of “gradient” concept might be more useful? And not a gradient lying on a single line, that would be too simple… but a big 3D gradient of connected nodes.