gamma-Aminobutyric Acid, otherwise more commonly known by its acronym, “GABA,” is a primary neurotransmitter of the central nervous system. While most of us may be more familiar with the concept of GABA as an inhibitory neurotransmitter produced by interneurons, helping to quell and refine activity of the excitatory glutamatergic pyramidal neurons, fewer people realize that it can itself be excitatory within certain cell populations.
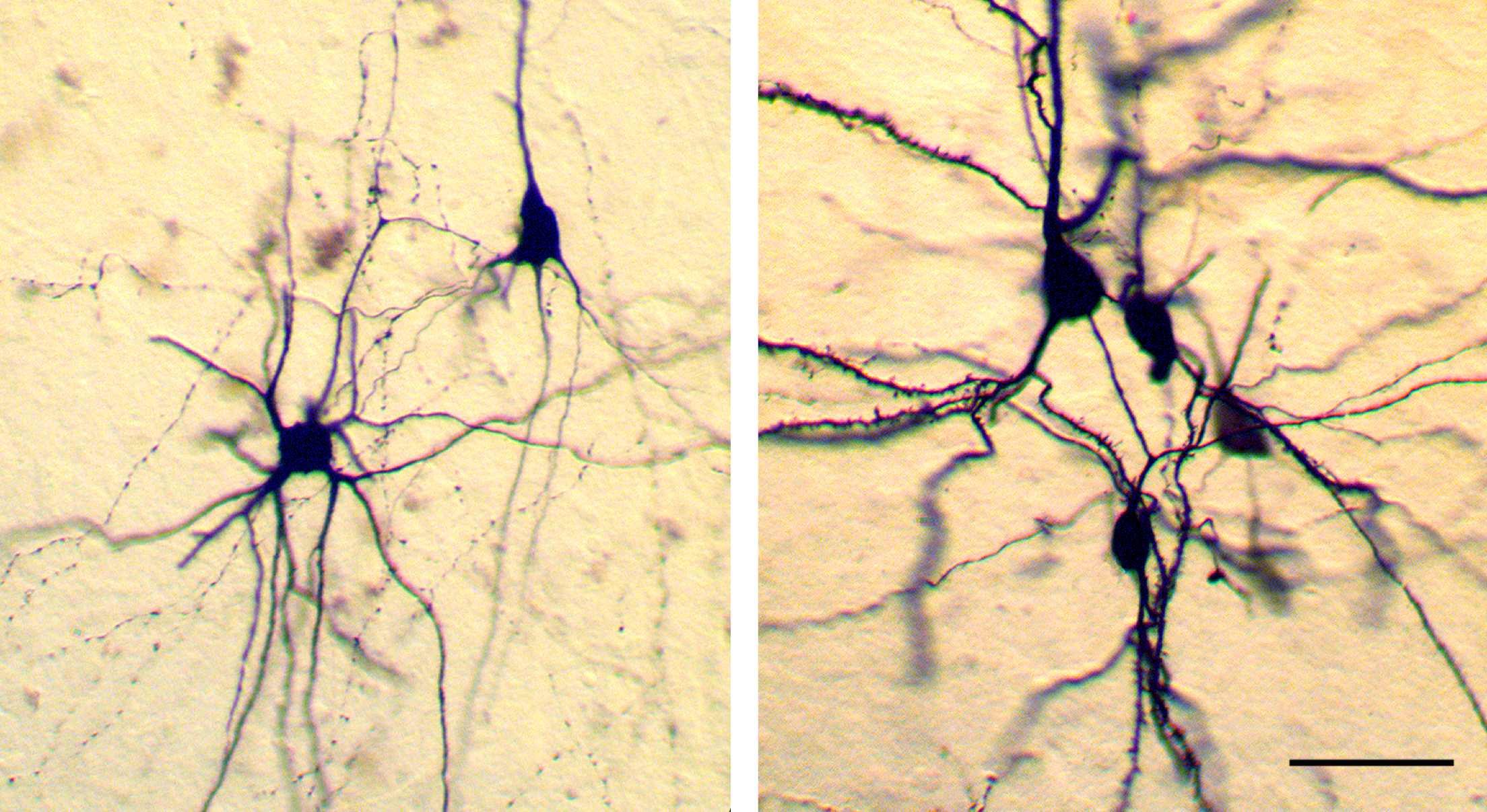
Some images of glutamatergic pyramidal neurons (triangular cell bodies) surrounded by GABAergic interneurons in mature cortex. Borrowed from here.
The primary reason GABA can behave as both an inhibitory and excitatory neurotransmitter is largely due to the combined expression of two chloride pumps and how much chloride, a negatively charged ion, is present within the neuron versus without. In the mature pyramidal neuron, mRNA expression of the chloride exporter, KCC2, is upregulated, meanwhile mRNA expression of the chloride importer, NKCC1, is downregulated. Thus, when GABA is released by a nearby interneuron and docks on a GABA receptor of a pyramidal neuron, the channel opens and chloride diffuses away from its higher concentration, which at this point lies outside the cell. This produces a rise in negative charge within the excitatory neuron, suppressing its activation. This is how GABA can behave as an inhibitory neurotransmitter.
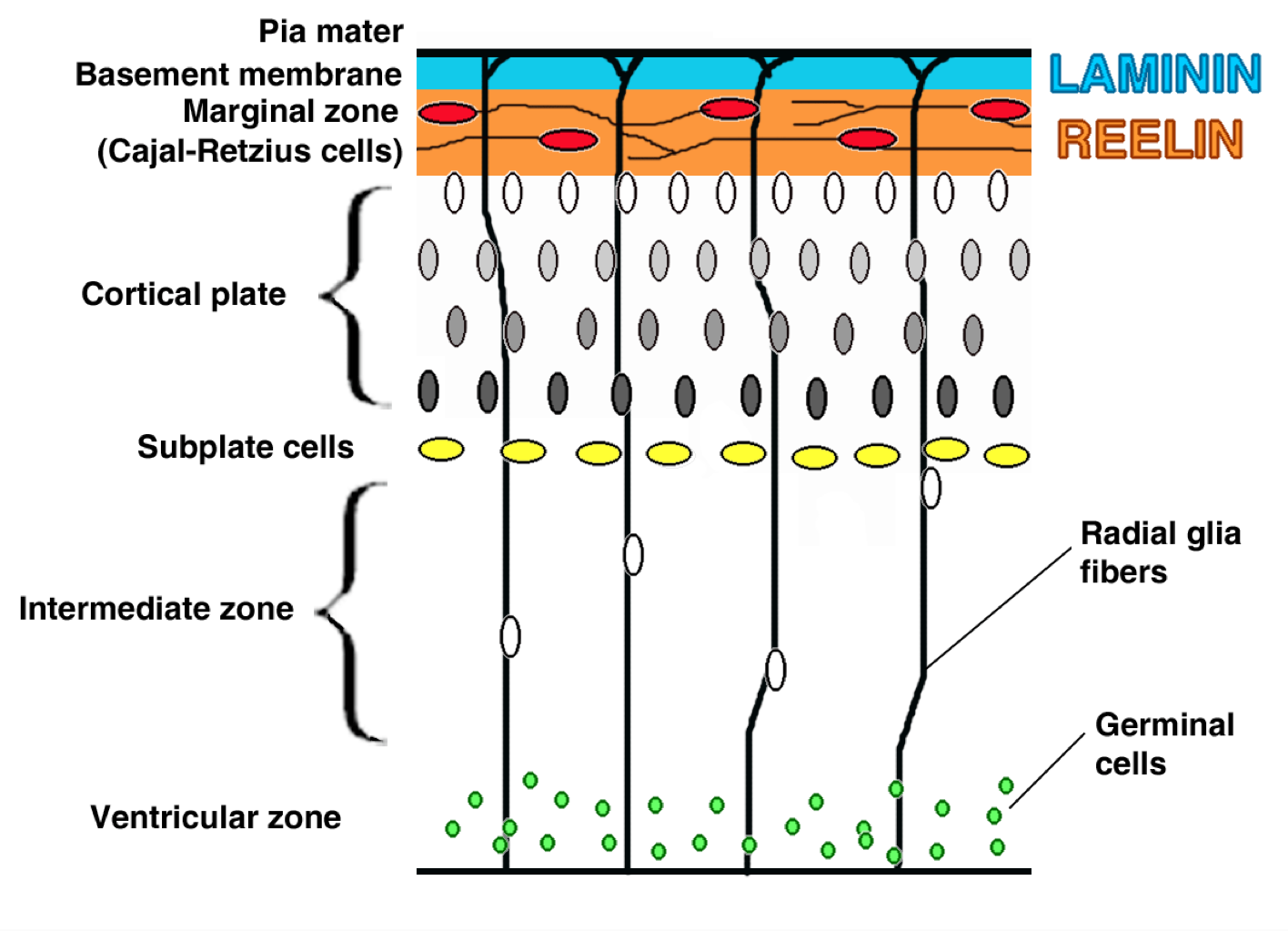
Interestingly, however, this is not the case with many types of germinal, newborn neurons (neuroblasts), and Cajal-Retzius cells. Each of these cell types express GABA receptors and also comparatively higher levels of NKCC1 and lower levels of KCC2, resulting in higher levels of chloride within the cells during a resting state. Thus, when GABA docks on a receptor in one of these cell types, chloride rushes out of the cell, increasing the positive charge within and potentially leading to excitation.
Thus, GABA is a vital driver of excitation in these cell types, which is most apparent during embryogenesis due to their large numbers, although can still be seen postnatally in the adult hippocampus which is continually regenerating portions of its dendate gyrus [1]. Excitation within progenitor populations tends to induce proliferation, either symmetric, in which more progenitors are produced, or asymmetric, in which two different daughter cells (e.g., a progenitor and a neuroblast) result. Excitation in general is integral in all processes of growth and development in the brain, and GABA is an early excitatory driver, largely because GABAergic interneurons are some of the first neurons to populate the developing neocortex.
Expression of NKCC1 could be considered a marker of neuronal immaturity and some cases of drug-resistant temporal lobe epilepsy lend support for this. Upon resection of the epileptic tissue in these cases, scientists have noted that mRNA expression levels of NKCC1 are often upregulated and KCC2 downregulated, suggesting that when GABA is released in these tissues, it is behaving as an excitatory neurotransmitter, not an inhibitory one, leading to the seizures from which these people suffer [2].
I can’t yet find indications in the literature that suggest that early interneurons synapse on radial glia (germinal cells) or Cajal-Retzius cells, although numerous interneurons do populate the same Layer I as do Cajal-Retzius cells for a time, suggesting that they may directly physically interact. Later in development, these Cajal-Retzius cells and GABAergic interneurons die off, leaving Layer I comparatively cell-poor. But interneurons do not populate the germinal zone where the radial glia are located. It is possible that interneurons could synapse on the long extensions that radial glia maintain, connecting them to the pial surface. However, some research suggests that GABA may also act in a paracrine fashion, meaning that it could be diffused in a non-specific manner into the extracellular space, reaching more distant targets than is typically seen in focalized synapses [3]. As such, it makes me wonder whether these interneurons, in a paracrine fashion, are driving excitation in radial glia and neuroblasts, promoting proliferation and maturation.
Ultimately, although we generally think of GABA as an inhibitory neurotransmitter, its action is context-dependent. For mature pyramidal neurons which have developed properly, yes, it’s inhibitory and allows the brain to refine excitatory signals. But in immature cell types, such as progenitors, newborn neurons, and Cajal-Retzius cells, it appears that GABA is an excitatory neurotransmitter instead. In fact, because it’s present within so many populations, including progenitors, I would consider it THE prototypical excitatory neurotransmitter, which evolution has further refined via a few switches of chloride pumps, broadening its usefulness. It makes me wonder if, at one point in evolution, GABA was the sole excitatory transmitter. Looking at the ages of the various chloride pumps may help to answer that question in future.

{kind=link}