Calcium ions (Ca2+) are the most widely used signal transduction molecules in living organisms, ranging from prokaryotes to complex eukaryotes such as ourselves. Calcium levels must be tightly maintained within the cell and are generally kept about 20,000 times lower intracellularly compared to the extracellular medium. On the other hand, levels of the ion, magnesium (Mg2+), are not quite as tightly regulated inside the cell. This is most likely due to the fact that calcium ions precipitate phosphates (e.g., ATP, ADP, GTP, etc.) which are the energy currency of the cell and would lead to considerable disruption in cell metabolism were they allowed freer reign. In fact, it has been shown that free calcium ions likely travel no further than 0.1-0.5 micrometers or 50 microseconds before they encounter some sort of binding protein, so there are a huge number of calcium buffers constantly present inside the cell [1].
It is believed that during evolution single-celled organisms initially developed binding proteins that acted as calcium buffers, sequestering it to certain locations within the cell or pumping calcium out entirely. Later on, prokaryotes probably began to take advantage of the presence of calcium as a signaling molecule, allowing the ion to bind to trigger molecules which in turn would effect enzymes or ion channels, propagating chains of reactions [1].
An excellent example of the exaptation of calcium for signal transduction involves the action potential of a neuron. In the typical neuron, the initial excitation is driven by the influx of positively-charged sodium ions into the cell. This then prompts the opening of calcium channels, a subsequent influx of calcium, further action potentials, and is typically associated with the presynaptic release of neurotransmitters or neuromodulators and their postsynaptic receipt by a neighboring neuron. The receipt of neurotransmitters/-modulators across the synapse can then lead to a whole new action potential in the neighboring cell, continuing the chain of action.
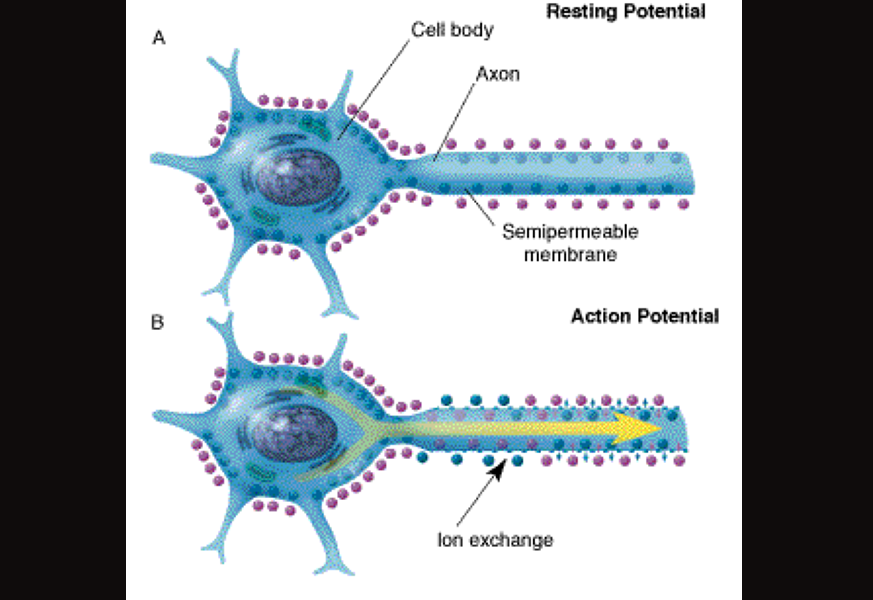
Illustration of the propagation of an action potential. Borrowed from here.
Even though we typically associate neuronal excitation with “learning,” most people don’t realize that the entire lifecycle of the brain is “activity-dependent”. For instance, the proliferation of neural progenitor cells (NPC) are dependent on the continual propagation of calcium waves between progenitors. These NPCs are directly connected by gap junctions that allow the free flow of calcium ions from cell to cell [2]. Similar calcium waves can be seen later in adulthood in the glial cells known as astrocytes [3]. When gap junctions are disabled in NPCs, neurogenesis or the production of neurons is suppressed. It appears as though certain baseline calcium waves in NPCs lower the threshold at which certain calcium-sensitive neurogenic transcription factors can be activated. Even minor suppression to these calcium waves can increase the threshold level at which NPCs respond to neurogenic signals such as various growth factors.
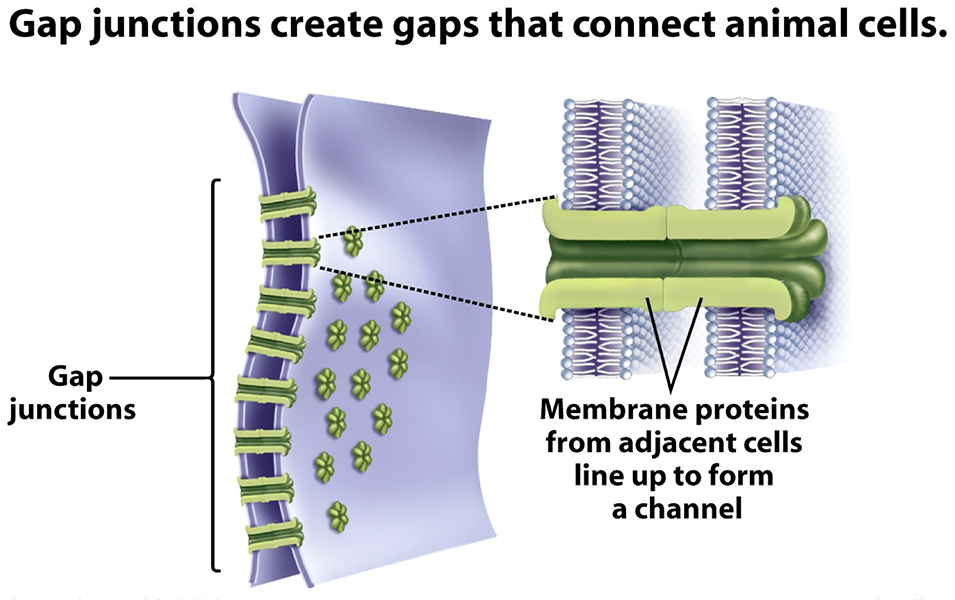
Illustration of gap junctions. Borrowed from here.
Neurons form branches or arbors, like a tree, which generally synapse on other cells, allowing cross-communication. The shape or “morphology” of those branches can be altered by changes in calcium signaling, affecting the length and overall complexity of the arbors and ultimately affecting how they communicate. Various neuromodulators, such as serotonin and acetylcholine, can also affect intracellular calcium signaling, leading to changes in branching complexity [4].
Finally, calcium affects learning through its vital role in longterm potentiation (LTP) or the strengthening of a synaptic connection between two cells through synchronous stimulation. This process is dependent upon the enzyme, CaMKII, whose activity is induced by calcium influx. Animals deficient in CaMKII fail to produce LTPs [5].
These are just some basics about calcium, and by no means do these examples summarize calcium’s many functions in the cell. They are simply examples to show you how calcium plays vital roles in the production of neurons, their migrations and overall shapes, and ultimately, in their behaviors. In short, neurons that are produced (if they are produced at all, that is) under calcium-deficient or -exorbitant conditions invariably show disturbances to their morphologies and to their behaviors, which vary by the specific conditions under which they arose. Some neurons may fail to have migrated out of the proliferative zone and into the cortex; other neurons may have migrated to the wrong places; others still may have made it to their target layer in the neocortex but exhibit disturbances in branching; and finally, some may exhibit differences in the number of synapses formed or how those synapses ultimately function.
Calcium & Neurodevelopmental Conditions
Conditions such as autism, epilepsy, schizophrenia, and bipolar have each had reported cases involving genetic mutations in calcium-related genes [6, 7]. The family of voltage-gated calcium channel genes are frequent favorites of mutation, perhaps in part due to large repetitive DNA content leading to relative genomic instability [8]. Mutations in CACNA1C in particular have been noted in each of these neurodevelopmental conditions. In relation to autism, although its mutation underlies the condition known as Timothy Syndrome which is itself extremely rare, its comorbidity with autism is the highest of all syndromic forms, quoted at over 90% concordance.
How do these different conditions all potentially share mutations in the same gene? Excellent question, one which currently has no definitive answer. However, to begin to unravel the mystery, it’s important to understand that these voltage-gated calcium channel genes produce MANY MANY MANY transcript variants– in other words, a single gene can produce many different gene products [9]. Which means that, although it’s the same gene, the specific mutations may differ between these different conditions, resulting in different phenotypes and behavioral outcomes. It’s entirely possible that the variation in gene expression may target different stages of cellular development or different parts of the cell. Currently, there’s considerable evidence to suggest that the stage of neurogenesis is targeted in autism and epilepsy. But in schizophrenia or bipolar, perhaps that may not be the case. Who knows? Calcium performs many divergent functions in the cell, and perhaps what we recognize as autism, epilepsy, schizophrenia, and bipolar are four sides of the same coin, each affected in slightly different ways, leading to different behaviors.
Time and further research will tell.

Hi Emily, you may want to check this website/ online publication ‘Central Role of Voltage Gated Calcium Channels and Intercellular Calcium Homeostasis in Autism’
http://autismcalciumchannelopathy.com/
Thanks, Natasa. 🙂 I think you had (maybe?) linked to that or a similar article before, and actually in writing this piece I was partially thinking of some of our previous conversations. Considering that calcium is at the heart of neurogenesis and neuronal differentiation in almost innumerable number of ways, the idea that it plays a fundamental role in autism development (as well as probably other neurodevelopmental conditions like schizophrenia and bipolar, as well as probably even Alzheimer’s and other neurodegenerative conditions) makes considerable sense. I’m currently working on a publication that involves the concept of “activity-dependent development” in neural development in autism, which of course naturally implicates calcium either directly or downstream of other calcium-inducing pathways, so I’m definitely right there with ya. Thanks again for the reference!
Pingback: Cold Snap: the physics of food security | DR FIONA McMILLAN - SCIENCE WRITER·