Okay, I’ll admit, the study of neurite (i.e., axon and dendrite) formation isn’t my area of expertise. My background is more in neuropathology and genetics. But I had been working on some revisions for an article now in press, and the reviewers wanted my neuropath point-of-view to be expanded a bit to include information about neural networks in autism. So, being an author who wants her paper published, I said, “Okay!” and skipped along to Google Scholar and PubMed to find more info on neuritogenesis in autism to supplement the paper.
Here’s a little background necessary to understand the point I’m about to make. A neuron, in its youth, produces branches called “neurites”. The process first involves a pretty straightforward elongation, followed by a switch to branching, kinda like a tree. In the adult neuron, these neurites can be subdivided into two general groups: the axon and the dendrites. If we’re talking about a neuron situated in the cortex, then it’s axonal branch will extend downwards, meanwhile the dendrites extend up and out. Depending on the cell type, the axon can shoot down into the subcortex, it can travel across the midline into the other hemisphere, or it can extend an axon and synapse somewhere else either nearby or far away within the same hemisphere. The axon is primarily the extension used for output, so that it can communicate with other cells. Dendrites, on the other hand, receive input from other cells over much shorter distances. Dendrites, except in rare instances of pathology, are not myelinated, which is a reflection of the shorter distances they traverse. (The longer the axon, the heavier the myelination usually.)
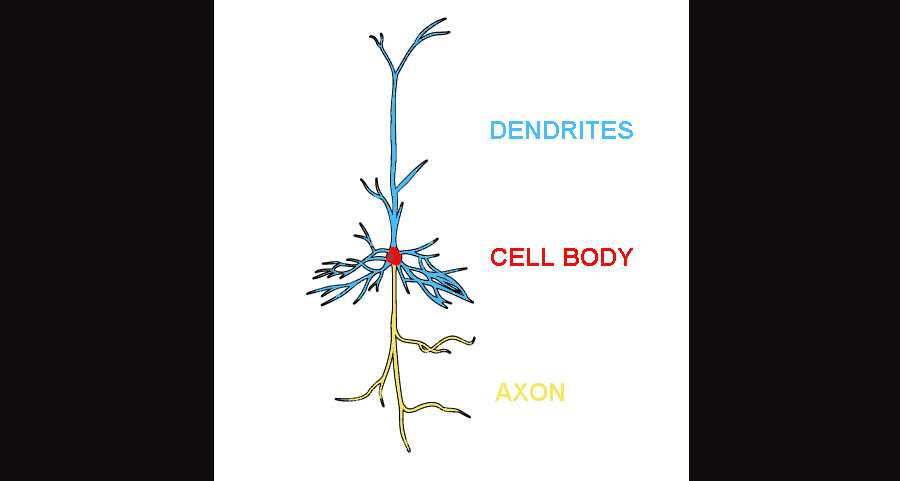
This is an illustration of the protypical layer V pyramidal neuron. The axon in yellow extends subcortically. The dendrites can extend the height of the neocortex, synapsing on various targets, including the cell-poor, synapse-rich layer I.
Anywho, in the life of a newborn neuron, the axon starts developing first in a process of elongation. Elongation of dendrites may partly overlap with this process but their developments are delayed compared to the axon. Both sets of neurites first elongate, and then branch. Elongation and branching are considered separate though interrelated processes. For the most part, elongation is a pretty permanent process: once the neurite ends up where it ends up, it either stays there, or can be pruned. But there’s usually very little opportunity for regrowing an entire arm. However, branching is usually a fairly plastic process and changes throughout the lifespan. Small branches can be formed, pruned, and reformed if need be. Axons, well, can’t.
In autism, MRI studies have reported a range of findings in autism; however, overall, the general theme is one of long-range underconnectivity and possibly short-range overconnectivity [1]. I’d like to point out, however, that these findings say nothing about unmyelinated extensions, which include all dendrites and some axons, because the resolution of MRI is currently too poor to detect anything smaller. Therefore, when we talk about long-range underconnectivity/short-range overconnectivity in autism, we’re talking solely about axons that are large enough to be detected.
So, because the numerous MRI studies suggest that axonal development is affected in autism, you’d think that there’d be huge numbers of molecular and animal studies investigating axon development, right?
Wrong.
If you go onto Google Scholar and type in the keywords “autism” and “axon”, plenty of page results are returned. However, if you start looking through the articles, axons seem to receive only a passing mention. Most often, the term seems to be lumped in with dendrititic formation or other features like migration. In fact, contained within the first 20 pages of the search, I found only 9 (out of 200) that specifically focus on axonal formation in models of autism, imaging studies aside.
Now, the study of neuritogenesis is not my area of expertise, I’ll admit. But even as a noob, I am shocked that there appears to have been so little study of axon formation in autism models given the wealth of human MRI evidence indicating that the axonal compartment is affected. I mean, “underconnectivity” is a very popular theory for autism, and yet the neuritic compartment that underlies the MRI findings (i.e., axons) has been overlooked. It seems instead that theory on neurite formation has been generalized and that the dendritic compartment has instead received far more attention. After all, let’s face it, “dendrites and synapses” have been the hot topic in autism for some time now. And I’m afraid that scientists might be forgetting (or perhaps aren’t even aware) that even though the developments of axons and dendrites have much in common with each other, they are technically separate neuritic processes with a divergence in the molecular factors that control them. And the same can be said of elongation and branching: they are distinct stages of neuritogenesis.
Not being an expert in neuritogenesis, I wonder if part of this trend is a reflection of technical limitations. For one, I would imagine it’s simply easier to study the dendrites than it is the axon, particularly if one is studying neurons in a petri dish or just staring at small chunks of tissue through a microscope. Dendrites are short processes and make up the bulk of branch formations, whereas in vivo intrahemispheric axons can span up to several inches in human. Even in mouse brain, the homologous tracts would still probably range in centimeters and be a challenge to study under a microscope. In a petri dish too it’s a challenge identifying precisely which neurite is the axon. Some studies simply label it by chronology: the first branch to develop is the axon. Other techniques may involve labeling proteins specific to the axon hillock or other axonal segments.
I also wonder if it’s a reflection of the division between fields of expertise. The underconnectivity/overconnectivity theory is based on imaging studies, whereas dendrites and synapses are the pets of the developmental and molecular biologists. A similar division can been seen between autism geneticists and neuropathologists (the latter of whom are few in number): genetics love the dendrite and synapse whereas the neuropathologists are more interested in neurogenesis. Neither group seems too interested in what the other is doing, with occasional exceptions, and thus there’s a theoretical barrier between the fields, leaving them poorly integrated.
In any case, I have to say I was genuinely surprised when I went searching for information on axon formation in autism models. For everything I’ve read or browsed through concerning the dendrite and synapse in autism, the field is comparatively barren when it comes to axons. But for whatever reasons, be they popularity, technical limitations, etc., hopefully that trend will change in future. If axonal formation and function is at the root of some of the core behaviors in autism, such as poor integration of social and language cues, it’s imperative that we try to understand how that occurs.
