A few weeks ago I summarized the findings of our latest study in Frontiers. Unfortunately, I gather as there was almost no interest in the blog that I did a very poor job of explaining why our findings are relevant to the current science of autism. My bad. To rectify that little blunder, I’m using this Sunday’s blog to break down our latest findings and explain why they’re important in continuing the work to understand what autism is at the cellular level.
Science is often a popularity contest, as are most other things in life. As such, the ideas en vogue may be a reflection of their usefulness, but may especially be indicative of which scientists and organizations are “on top” in the hierarchy of scientific competition. This is nothing new to the field. This also means that certain ideas, whether accurate or not, may dominate a field of research for a period of time. Just look at the Refrigerator Mother Theory of autism from the 20th century.
If one has attended the International Meeting for Autism Research for the last several years running, one quickly gets the impression that the bulk of the scientific community has decided that the pathology of autism lies within the synapse. This, my friends, is the hot topic in autism. And it’s not to say that there isn’t strong evidence supporting the role of synaptic dysfunction in the heterogeneous conditions. However, one thing which the bulk of the community seems to be failing to do, in my estimation, is view the development of the neuron as a type of gestalt. Instead, the trends are to dissect portions of the neuron, such as the synapse or dendrite, and treat them as though they’re anatomically separable from the remainder of the cell. As such, we only get photographic glimpses into the life of that cell, which do little to tell us how it developed in the first place.


Work by various neuropathologists, including my husband, have shown that over 90% of brains studied postmortem in autism exhibit disturbances to early neuronal development, such as neurogenesis and pre-migratory maturation. This is particularly apparent in the neocortex and is evidenced by dysplastic structures, heterotopias, etc. In spite of this evidence, the topic is not currently popular and therefore doesn’t typically factor into most theories of autism. Reflective of this, if one were to keyword “autism” + “synapse” into PubMed, you would receive a return of 653 articles, compared to “autism” + “neurogenesis” which returns only 157. Do the same on Google Scholar, and “autism” + “synapse” returns over 20,000 hits, meanwhile “autism” + “neurogenesis” only 12,000.
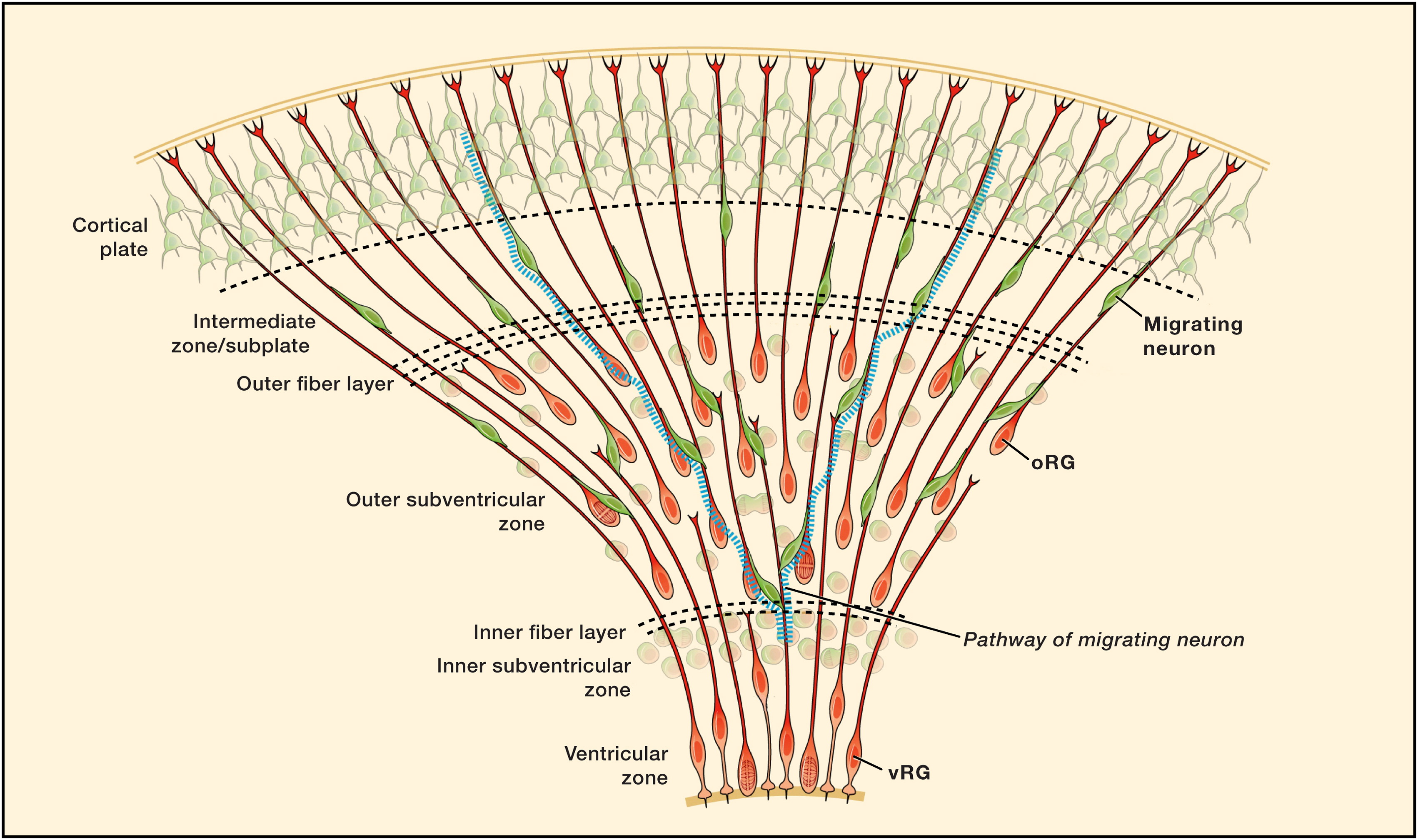
Illustration of neurogenesis in the developing neocortex of the human brain. Newborn neurons are produced from the ventricular and subventricular zones and then migrate to their final destinations in the cortical plate. Borrowed from Lui et al. (2011).
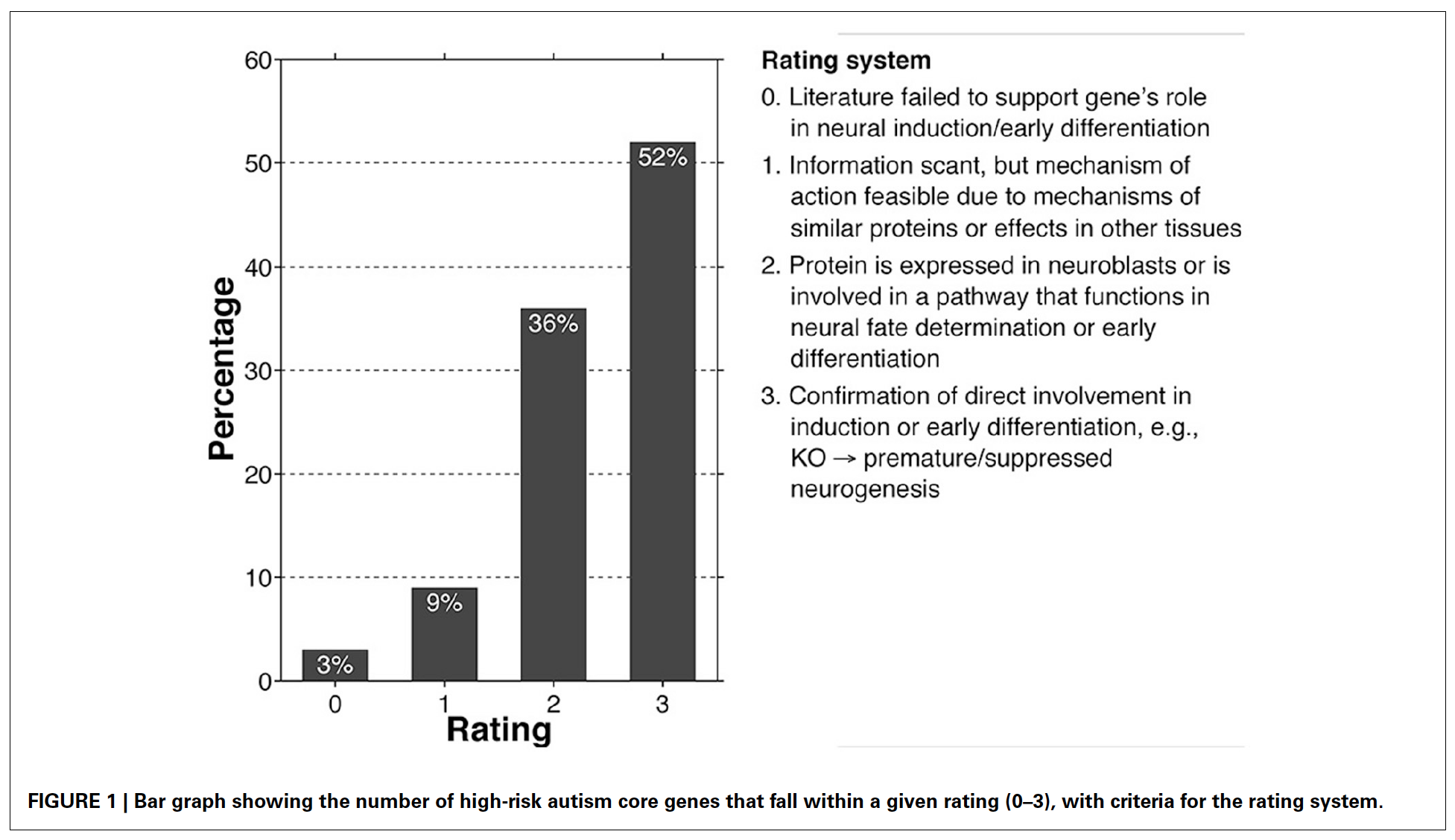
In this last publication, our greatest concern was rectifying these divergent evidences with one another. In order to understand whether both the neuropathologists and molecular biologists are correct, we took a list of close to 200 high-risk autism-related genes and delved in depth into their involvements at different stages of neuronal development. What we found was that 88% of this core list of genes showed involvement in early stages of neuronal maturation, ranging from neural induction just preceding neurogenesis to pre-migration. Most often this manifest as premature or delayed neurogenesis. However, 80% of these same genes also appeared to influence later stages of post-migratory neuronal differentiation, in terms of neuritic and synaptic developments as well as ongoing plasticity.
In short, this work suggests that the genes implicated in autism tend to affect multiple stages of neuronal development and it’s therefore important that we study development of the neuron in autism more holistically. (We should also study cells other than neurons as well, but that’s a topic for another blog.) This work also indicates that both neuropathologists and molecular biologists are correct, albeit in looking at and describing different sides of the same coin. We’re hoping that this work is a positive step towards integrating these somewhat disparate fields of research.
While I would be one of the first people to admit that the behavioral symptomology of autism is largely due to how the brain connects and communicates within itself, I also believe that many aspects of neuritic and synaptic developments are rooted in earlier processes of neuronal maturation. Therefore, even if we wish to study the synapse in autism, we may still need to look earlier in the neuron’s history in order to do so. In metaphorical terms, I prefer to view the life of a neuron as a long string of molecular dominoes: even when just one domino is out of place, the development of that neuron can be irrevocably altered.

Hi Emily,
Regarding your earlier and current post on this research, it’s “my bad not your bad” for rarely getting it together to reply with the huge amount of enthusiasm and questions that expert and thoughtful posts generate. True to form, I’m wimping out again right now. My reasons are that I’m about to go on a group hike with my son and that I’m really bad at keeping up with life in general.
It’s much appreciated when scientists take the time to share ideas with a broad audience!
Erica Eaton
Hey, no worries, Erica. Glad to know it’s piqued your interest. Hope that the update today is additionally helpful. Enjoy the hike. 🙂
Is one idea that it makes sense to look more broadly over the time course of development, especially because many high-risk autism-related genes appear to influence multipe stages of neurodevelopment? If I understand correctly, brain cells change form and move in synchronized patterns. If something goes awry at one point in time, the change will (of course) continue to influence the overall pattern as time goes on. I watched Manuel Casanova’s very interesting presentation “Heterochronic germinal cell divisions and abnormalities of migration in autism” to try to understand some of what you writing about a little better.
May I ask if some of the structural differences you mention have any relationship with/analogy to structural differences such as epicanthal folds or midface hypoplasia? Are the differences also associated with other neurodevelopmental or psychiatric disorders? Do they suggest anything about what might underlie Christopher Gillberg’s observations (described in SFARI link) of some association between significant language delay in early childhood and somewhat different issues in later childhood (e.g. ADHD, autism, etc.)? http://sfari.org/news-and-opinion/viewpoint/2013/essence-from-defined-categories-to-broad-understanding
Thank you very much to the authors for your research, blogs and presentations.
Hi, Erica. Yes, there’s definitely something to the idea that earlier stages of cell development lie foundational to later stages, something which tends to be overlooked. There is also evidence, however, that some of these gene products function at multiple time points in cellular development. Evolution really likes to recycle things rather than create anew, so quite a few of the genes we studied would also fit into that category, although a single gene usually provides multiple different spliced gene products known as “transcript variants” and therefore different transcript variants may be used at different time periods of maturation.
In short, the genes appear to influence most stages of neuronal development, either by being a foundational element in earlier neural development leading to downstream disruptions in later processes, or by performing functions at different time points of cell maturation. The interesting thing is that most of these genes perform functions which are integral to cell development. That might be a no-brainer, but when one considers what that means, it means that these gene products are produced in order to INDUCE CHANGE in a cell line. To change to some other type of cell, usually from some sort of stem cell or progenitor line. This is in contrast to gene products that are involved in homeostasis and help to keep the cell going as it is. Thus with autism, it suggests that there are disruptions to cellular change, otherwise known as “differentiation”. In this publication, however, we elected to use the term “maturation” to broaden the idea to include earlier stages which most cell biologists would consider as part of “neurogenesis” but which are also part of that process of change. “Differentiation” tends to be thought of as later changes in cell maturation, such as synapse development, etc. Does that make more sense?
Yes, it does! That’s interesting that there may be a tendency for autism-relevant genes to be especially involved in change (vs. homeostatis). It’s interesting, too, about how the word “differentiation” is used in practice, and how you chose to use the word “maturation” to help broaden thinking to include relatively early changes.
The idea that “some of these gene products function at multiple time points in cellular development” also came through. It’s fun learning about how things are “recycled.” By the way, while I do know that genes are turned on and off over time, may I ask about the gene products known as “transcript variants”? Do you mean that DNA contains a bunch of genes that are variants on a theme, or instead that sometimes only part of an individual gene’s code is used, resulting in protein variants that are made from “subsequences” of the gene?
If you have time, I have a couple of other questions that probably aren’t as relevant to the blog. If it’s OK, I’ll email you at scienceoveracuppa@gmail.
Sure, go right ahead and send me an email.
To answer your question, a single gene is generally used as a blueprint to produce different transcripts, which are RNA. This RNA can then be used to produce protein (or the RNA itself may serve some other function and remain RNA). Basically, different transcripts are just different splices of the same gene product which ultimately can lead to different functions for those products. So, as a simple example, if you’re familiar with the idea that exons, which are the coding sequences of DNA, are usually separated by introns, which are usually thought of as non-coding elements. When a gene is transcribed, the whole thing is produced into RNA, introns and all, and later these introns are spliced out. Well, some introns or parts of introns, may only be cut out in certain circumstances, such as when Protein “X” binds to a portion of the RNA. So if Protein “X” binds, then the splicing machinery knows to cut out Intron “Y”. If Protein “X” is nowhere to be found, then the splicing machinery leaves Intron “Y” where it is and then Intron “Y” may actually become part of the coding sequence, ultimately changing the protein sequence and its function. This can also happen to sequences of RNA that come before or after the first and last exon, known as “untranslated regions” or UTRs. Some genes have HUGE numbers of transcript variants. It’s believed, for instance, that the gene Neurexin 1, which is one of the largest genes in the human genome at over 1,000,000 base pairs, may have over 1,000 different transcript variants. This is probably in part due to the fact it has HUGE introns which contain many different regulatory sequences that can bind different regulators and create subtle differences in transcript splicing.
Hi Emily, I would be interested to hear your take on this brand new paper and if their finding fit what you are observing (spotting the word ‘neuronal progenitors’ in this para made me think of you 🙂
“… These data provide support for a mechanistic connection for viral-infection hypotheses for autism with neural over-growth hypotheses through the novel identification of exaggerated M2 activation states in autism brain tissue.
…. We observe, for the first time, that M2-activation state microglia genes, in particular, are altered in autism, potentially driven by type I interferon responses. This process may drive changes in neural progenitor cell proliferation and connectivity with resultant altered activity-dependent neural expression profiles in post-natal development. The linkage of this pathway to autism may lead to more accurate and predictive models of idiopathic disease that might contribute to the identification of effective therapeutic approaches. …” http://www.nature.com/ncomms/2014/141210/ncomms6748/full/ncomms6748.html
Yeah, any kind of stress response in the embryonic brain I could see as a potential risk factor for autism. I haven’t read beyond the abstract of that paper, but just to mention that if they didn’t address the cause of death for the autism group, that could be a confounding factor in their study since they’re looking at gene expression rather than sequence. This is often an overlooked confound in these types of expression studies. For instance, quite a few autistic children have the misfortune to die from drowning, meanwhile few to no controls ever do. Many times this isn’t an immediate death by drowning, but they may be resuscitated, brought to a hospital, and die some time later. The issue with this is that drowning will definitely cause cellular stress and induce a whole immune response in the brain. Any death that isn’t immediate has the potential to change gene expression. Therefore, it can be difficult to be certain what researchers are actually measuring, the autism or the mode of death, in these types of expression studies. My husband, who’s more of an expert in this area, is constantly complaining about this overlooked confound.
But in any case, it would make sense if immune-related pathways were involved in some cases of autism risk. I’d definitely agree with that.
Thank you, Emily! Over the years, I’ve retained simple concepts such as that of “templates” (for lack of a better word), and that of a progression from DNA to RNA to protein. Thanks very much for a reminder of/introduction to multiple subtleties. In working on my understanding, I found this very brief video: http://www.dnalc.org/view/16932-Flow-of-Information-from-DNA-to-RNA-to-Protein.html
Great, glad to be of help. 🙂