Awhile back, I wrote about how the neurotransmitter, GABA, can behave as an excitatory signal in certain cell types rather than the inhibitory signal with which we’re more familiar. As a quick recap, a neuron generally fires when the internal charge becomes more positive and reaches a certain threshold voltage, a process known as depolarization. GABA is a neurotransmitter that docks on its receptors, opening the channels to allow the ion, chloride, into or out of the cell. Whether it comes in or goes out depends upon its concentration gradient, whether there is greater or lesser concentration of chloride within the cell. If chloride is greater outside the cell, such as occurs in most mature neurons, it will rush in and because it’s negatively charged, it will suppress neuronal excitation (i.e., the “action potential”). If, however, there is greater chloride concentration inside the cell, chloride will rush out, and may lead to an action potential if the neuron depolarizes enough to hit the threshold for firing. In this way, GABA can be either inhibitory or excitatory. Excitatory GABA signaling is a vital feature of the developing brain and without it, development goes terribly awry: neural stem cells don’t divide as they should, neurons aren’t produced properly, and those which are produced don’t develop correctly.
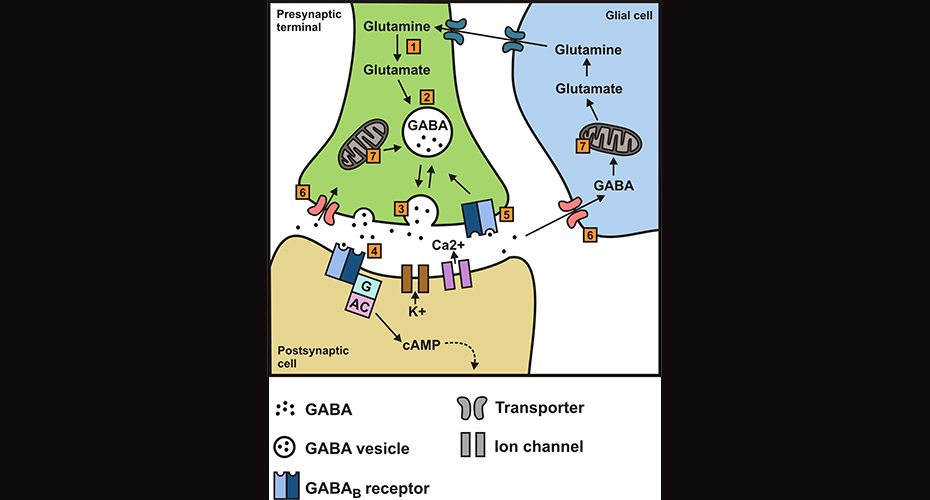
As I also discussed previously, the ratio of two chloride transporters are particularly vital in determining how a cell will react to GABA reception: NKCC1 and KCC2. NKCC1, if you’ll recall, is a chloride importer and is typically downregulated in mature neurons and upregulated in immature ones. Meanwhile, KCC2 is a chloride exporter and exhibits the opposite trend. Typically, in postnatal central nervous system, higher ratios of NKCC1:KCC2 are seen only in select cell populations such as the olfactory bulb, the hippocampus, the granular layer of the cerebellum, and the dorsal root ganglia [1].
Disturbances to the ratio of NKCC1:KCC2 have been reported in several neurodevelopmental conditions, including autism, epilepsy, schizophrenia, and bipolar. In some postmortem studies of people with these conditions, higher-than-usual ratios of NKCC1:KCC2 have been noted, suggesting the retention of an immature phenotype and disturbed cell differentiation. In addition, the drug, bumetanide, which specifically antagonizes the chloride importer, NKCC1, has found recent success in treating symptoms of both autism and epilepsy. In particular, Yehezkel Ben-Ari’s group has spear-headed much of the work on excitatory GABA in autism and has been involved in some of the early drug trials of bumetanide.
Of relevance to this line of research, I was recently reading an article published by Damborsky and Winzer-Serhan (2012) that suggests that the switch from excitatory to inhibitory GABA during very early development may vary in mammals according to sex. In short, what the authors found was that by postnatal day 5, female rat pups had lower NKCC1:KCC2 ratios as compared to male siblings, suggesting that females exhibit a more mature phenotype than males by this stage.
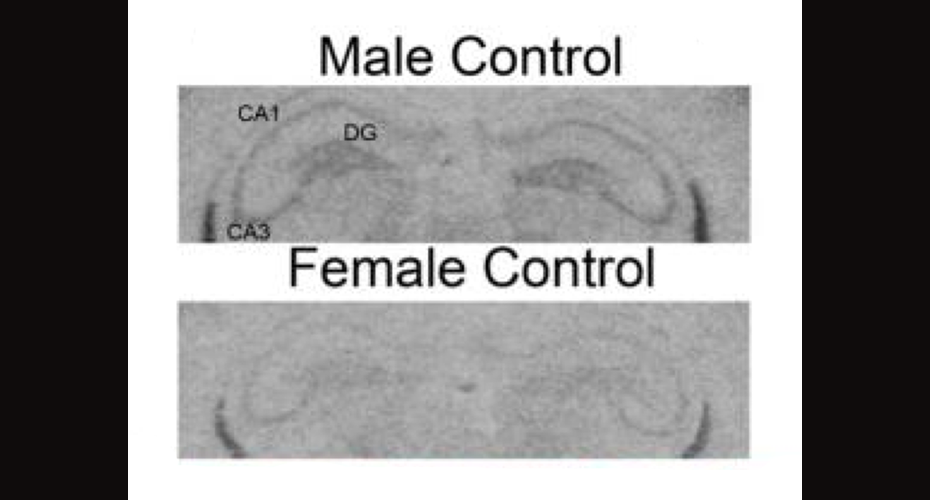
A section of Figure 2 borrowed from Damborsky & Winzer-Serhan (2012), showing an example of differential NKCC1 mRNA expression in hippocampus in male versus female control.
In addition, because nicotinic receptor activation is associated with the developmental switch from NKCC1 to KCC2, the researchers went on to study the effects of application of nicotine (via ingestion) on NKCC1:KCC2 ratios. As expected, early postnatal application of nicotine led to an earlier drop in NKCC1:KCC2 ratios in males in hippocampus– although curiously this effect was not seen in the females. By P8, these effects were no longer significant even in males, indicating a brief time window of vulnerability.
So, why am I so interested in this study on GABA excitation in rat pups? Well, considering the lop-sided sex ratios in autism, with males receiving diagnosis far more often than females, this research suggests that males could have an extended time-window of vulnerability as compared to females, if indeed GABA excitation plays some role in the development of autism symptoms and is not just a secondary symptom in postnatal life. That’s not to suggest that this is the ONLY factor underlying differences in risk according to sex in autism. It may, however, be an important one and worthy of further study.
