What is a microexon? The explanation may be slightly complex for those not familiar with the basics of genetics, so I’ll review a bit. First off, a gene is a section of DNA that is usually comprised of certain functional segments. This includes a promoter region at the start of the gene that binds with various proteins and RNA (usually known as “transcription factors”) in order to kick off gene expression, hence the name “promoter”. After this there’s usually a collection of alternating exons and introns. Exons are portions of the gene that are used to produce the final gene product, be it protein or RNA. Introns, on the other hand, are usually cut out following transcription. Then we have microexons, which are short segments of DNA, usually 3-27 nucleotides in length, that are essentially alternative exons: sometimes they’re spliced out, other times they’re included in the final transcript. Variability in splicing events is known as alternative splicing (AS) and takes its cues from the different binding partners that may latch onto the transcript.
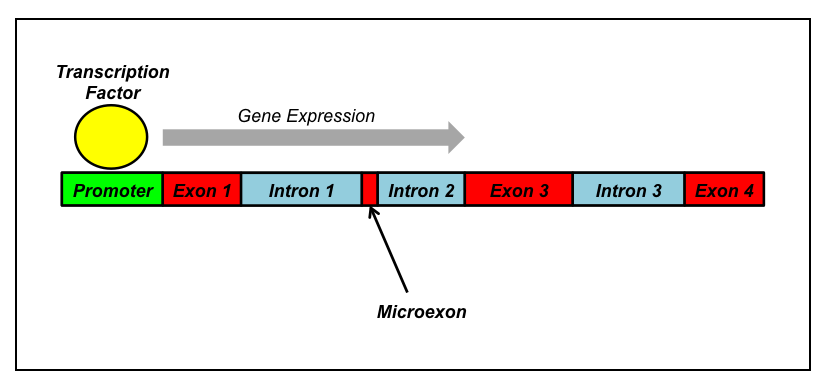
Most typically, the inclusion of microexons in a transcript is associated with the extent of differentiation (maturation) of that cell line. This is especially true for neural cells. Thus, for instance, even though a neural stem cell and some of its progeny may both express a particular protein, let’s called it Protein X, the neural stem cell is less likely to produce Protein X that contains the portion coded by the microexon; meanwhile its neuronal offspring produce Protein X that includes the sequence coded by the microexon. Scientists believe that this type of microexon expression is likely associated with changes to the function of the gene products these sequences are embedded in and plays an important role in cell identity and cellular maturation. Microexons are likely important in instructing neural stem cells to undergo neurogenesis and continue further differentiation. In support of this, when included in the final gene product, microexon sequences are usually within an easily accessible portion of the protein, suggesting their availability to and interaction with other binding partners. In particular, microexon sequences are often very near portions of the protein which fold, indicating that interaction with a microexon sequence can alter the shape of the nearby folded region and change the overall shape– and, therefore, function– of the protein itself. Shape, after all, defines function.
Relevance of Microexons to Neural Development
The recent article in Cell by Irimia et al. (2014) has shown that, more than any other cell type studied, neuronal development is especially subject to microexon-related AS events. Other longer splicing events were not as significantly related to neuronal development. Most surprisingly, although microexon events comprise only about 1% of total AS occurrences, they made up approximately 1/3rd of neural-related AS events. In addition, these microexons are often highly conserved across many species, spanning back hundreds of millions of years. The intronic regions surrounding the microexons also show stronger cross-species conservation, indicating their importance to very basic mechanisms of neuronal development.
The researchers found that increased microexon inclusion occurred at its height in mouse brain after neurogenesis, during a time when newborn neurons are still developing, e.g., migration, neurite sprouting, synapse development, etc.
Relevance to Autism?
Irimia et al. used 22 samples from the brains of autistic individuals and 20 controls. They then looked at RNA expression, determining that on average autistic brains had more microexons spliced as compared to control brains. This trend wasn’t consistent across all samples however, but only present in a portion of cases.
Unfortunately, as is the bane of many a neuropathologist, so infrequently do scientists take into account factors such as modes of death when it comes to gene expression studies. Nothing is mentioned in the article as to how these people died. One thing, for instance, which is all too common amongst autistic deaths but is rarely seen in controls is drowning. Many times, the individual may not die immediately but is resuscitated for a period of time before succumbing. This can often lead to serious confounds in gene expression due to the changes hypoxia and cell stress inflicts on the brain. The other shortcoming of gene expression studies is that they are dealing with gene expression postnatally, not pre- or perinatally. Often, the developmental trajectory can change considerably over time and therefore we must be exceedingly cautious to presume that the clues we glimpse now hold answers to how the condition arose in the first place.
Though this study is extremely interesting, the section on autism leaves more questions in my mind than answers. We can say that microexon expression is abnormally upregulated in a portion of autistic cases at the time of death compared to the control, but we don’t know the reasons. For one thing, it is acknowledged that cell stress responses often induce considerable changes to alternative splicing events [1]. Aside from confounds due to modes of death, comorbid conditions such as epilepsy often lead to considerable cell stress and may potentially change the transcriptomic landscape. It’s entirely possible that, because we are viewing positive findings in only a subset of autistic individuals, these results may be due to something entirely different than the original developmental cause(s) of the condition. Sadly, due to the sparse methodology and homogenization of data, we can’t tell.

Ah, reductionism. Quarks – could they unfold the true causes of autistic behaviour……..?
Quite possibly! 😉