If you’ve ever seen a dog follow a trail by scent, you’ll have gotten a glimpse of the importance of olfaction in mammals. Some scientists have postulated that while the sense of smell is not inordinately important to our reptilian cousins from whom placentals, monotremes, and marsupials arose, to mammals the nose has made all the difference. (Interestingly, geckos may be a notable example to the contrary [1].) It is believed that many early mammals found a nocturnal niche in which to avoid their diurnal dinosaurian predators,– though the strict diurnality of sauropsids is hotly debated and unlikely [2]. Nevertheless, it may be true that mammals were better at not getting eaten at night than at day. And for any nocturnal animal, vision is not usually its primary source for sensory information and it must instead rely on other senses, such as smell.
The earliest of early mammals had comparatively small olfactory bulbs. For those not familiar, in humans the olfactory nerve (cranial nerve I) and its bulbous tip lie at the underside of the brain, surrounding the midline of the frontal lobes. The circled portion in the image below indicates the bulb.
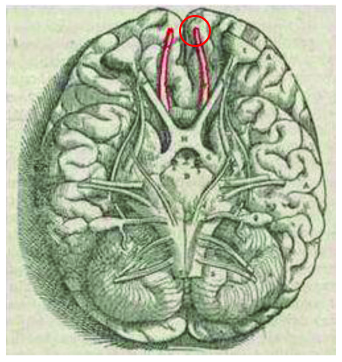
In other nonhuman modern mammals this bulb is often much larger compared to the size of the forebrain. Here for example is a superior view of a mouse brain, with the paired bulbs jutting out from beneath the frontal lobes:
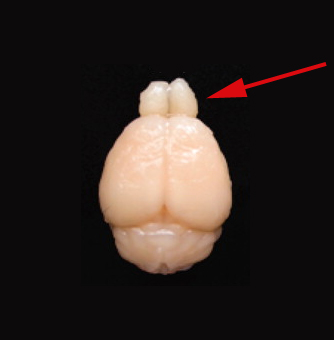
It is believed that early mammalian evolution involved three separate time periods of brain expansion, each one paired with the expansion of the olfactory bulb [3]. Rowe et al. (2011) postulate that the expansion of sensory modalities, e.g., olfaction, drove cerebral expansion. A hypothesis not without its charms.
However, it is also entirely possible that frontal lobe and olfactory bulb expansion grew together because they share similar ontogeny or developmental origins. While, as humans, we can easily see the adaptive benefits of sharper sensation and expanded frontal lobes, the relationship between brain and bulb merely confirms that there is a relationship and not what the original catalyst for their paired expansion may’ve been.
The mammalian brain is different from the reptilian brain in numerous ways, but when it comes to the cortex in particular, reptiles have only what would be considered the homologue of the three lower layers of the mammalian cortex. The mammalian cortex on the other hand has an expansion of layers, numbering six in total. The layers which most set mammals apart are layers II and III. In humans, these layers provide for local and long-distance lateral (side-to-side as opposed to down-up-down) communications between cells.
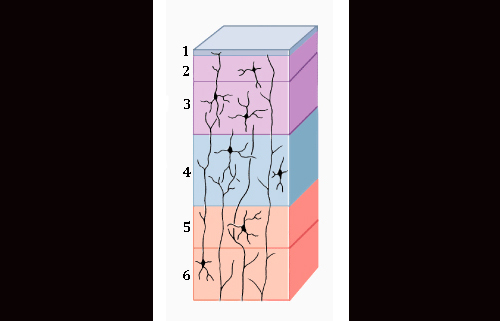
Most interestingly, the neurons from these layers are largely derived from the subventricular zone (SVZ), an area which lies in between the ventricular zone and the developing cortical plate and produces intermediate progenitor cells, a form of neural progenitor. This SVZ is believed to be the seat of radial (vertical) expansion of the mammalian cortex [4].
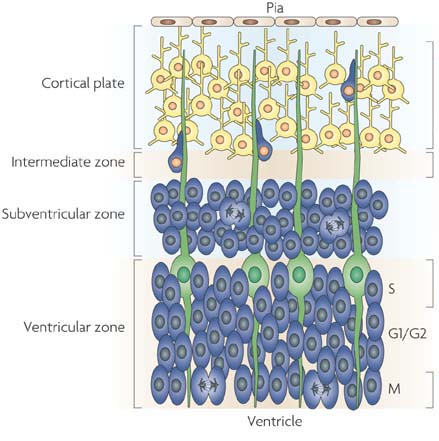
Now, would you be surprised if I told you that neurons for the olfactory bulb are also supplied by the SVZ?
“–Whoa, hang on…” you say to me. “Are you telling me that the same population of founder cells provides neurons for both the expanding mammalian cortex and the olfactory bulb?”
“Why, yes. Yes I am.”
“Holy shit that’s cool.”
“Yup. I know.”
Though they’re derived from different subpopulations of cells, arising from the same region they could both be targets of the same effectors. If Effector A targets the SVZ, it could result in paired changes in the cortex and olfactory bulb. Interestingly, one study by Yousem et al. (1996) looking at twenty-five cases of congenital anosmia (no sense of smell) or hyposmia (reduced sense of smell) in which the olfactory bulbs/tracts were either missing or poorly formed, 20% of those cases exhibited reduced frontal and temporal lobe volumes. Coincidence?
They say neurons of a feather flock together. Or something like that. And so far the three distinct expansions of the mammalian cortex and olfactory bulb suggest a significant relationship between the two. While most paleontologists may explain this as an adaptive result of change-in-one driving change-in-another, I instead see the possibility of two closely related results following a common cause.
And who knows what that common cause may be. Changes in food source, other changes in lifestyle, changes within the molecular environment we can’t even fathom, or all of the above. Whatever it was, the evidence, at least to me, doesn’t suggest that olfaction initially drove brain expansion or brain expansion drove olfaction– although that’s the easiest scenario to envision and a common one paleontology falls back on, the Natural Selection of Behavior. But sometimes environments just change, and with that change they bring adaptations in biology which need not initially be meted out at the level of inter- and intraspecies competition.
In the case of neuronal proliferation, proliferation had to increase before it could be selected for behaviorally. It may well have been reinforced through behavior, but that was not the first step towards expansion.
